

2016年04月の記事
全30件 (30件中 1-30件目)
1
-

興味津々、脱皮殻
大阪市立自然史博物館で開催されている特別展「生命大躍進」について、まだまだ書きたいことがあるのですが、いろいろネタがたまってきたので、とりあえずいったん休憩。さて以前に脱皮したオオコシオリエビ(Cervimunida princeps)のこと。熊野灘(深海)からやってきて初の脱皮でした。今回、脱皮殻を食べるか否かについて興味があり、脱皮殻をそのまま放置していました。海水の甲殻類は自分の脱皮殻を食べるもの、食べないものがいて、このあたりが個人的に面白い点です。例えばパープルリーフロブスターは自分の脱皮殻を比較的よく食べるほう。このオオコシオリエビはどうなのかと思い、毎日観察を続けていましたが、全く興味がないようです。次第にバラバラになっていく脱皮殻。でも誰も興味がないわけではありません。こちらはサガミモガニ(Pugettia sagamiensis)とヨコヤホンヤドカリ(Propagurus obtusifrons)。このうちヨコヤホンヤドカリがよく脱皮殻をハサミで挟んで、口元へ運んでいるところを見かけます。でも実際に食べているのかどうかはわかりませんけれどね。さてさて今度はヨコヤホンヤドカリが自分の脱皮殻を食べるのかどうかに興味が出てきました~(笑)。脱皮するようなことがあれば観察したいと思います。
2016年04月30日
-

大阪市立自然史博物館:特別展 生命大躍進 脊椎動物のたどった道
大阪市立自然史博物館で開催されている「特別展 生命大躍進 脊椎動物のたどった道」へ行ってきました。今回、特別展ブロガー招待に応募し、当選いたしました。光栄なことです。誠に感謝いたしております。特別展会場の入り口はネイチャーホールの2階です。貴重な化石の展示や最新の知見など、注目すべき点がたくさんあり、非常に楽しみでした。では早速~♪地球誕生はおよそ46億年前。この数億年後に生命が誕生したと考えられています。最初の生命誕生の痕跡が37億年前の岩石にありました。グリーンランドのイスアで発見された最古の生物の化石。この岩石には炭素(グラファイト:Graphitic Schist)が含まれており、解析の結果、炭素が生物由来であり、最初の生物の痕跡であると考えられています。(写真の黒い層が炭素の層。)こちらはオーストラリアの西オーストラリア州から発見された35億年前の微生物化石(層状チャート:banded chert))。チャートは二酸化ケイ素を主成分とする岩石で、岩石には微生物のかたちが残されているとのこと。そして27億年前のストロマトライト(Stromatolite)。オーストラリアの西オーストラリア州ピルバラ地方で発見されたもの。こちらは20億年前のストロマトライト。カナダのグレートスレーブ湖地域で発見されたもの。ストロマトライトはシアノバクテリアにより作られ、シアノバクテリアは光合成により地球の大気に酸素を作ったとされています。原始大気にはほとんど酸素がなく、酸素を必要としない生物にとって酸素は毒でした。現在酸素を必要としない、光合成に依存しない生物は深海の化学合成生物群集などに見られるのみで、大半の生物が酸素を必要としています。ちなみにシアノバクテリアは海水生体の飼育を行っているホムラspには馴染み深いバクテリアです。個人的に興味深く拝見させていただきました。とまぁ、すでに先カンブリア時代だけでも濃厚かつ貴重な化石の展示でした。ここから先は先カンブリア時代末期のエディアカラ生物群の展示です。でもその前に今回のテーマである生命大躍進として、注目すべきイベントが起こっています。それは約23億年前のこと。原核生物の中から真核生物が誕生しました。その後の多細胞生物へとつながる大躍進です。ではいよいよ全球凍結時代を含む、先カンブリア時代末期のエディアカラ紀(約6億3500万年~5億4100万年前)の生物群の展示です。このときこれまで見られなかった大きな生物が誕生しましたが、絶滅しており、古生代以降の生物とは類縁関係がはっきりしないとされています。遺伝子的に見ても、現在の動物が共通の祖先、共通の遺伝子から進化してきた点とは違い、異なる遺伝子を使っていたのかもしれないと推察されています。ではそのようなエディアカラ生物群の化石を紹介。アルボレア(Arborea arborea)。南オーストラリア産。現在のウミエラに似ている生物ですが、類縁関係ははっきりしないようです。ディッキンソニア(Dickinsonia)。南オーストラリア産。ロシアの白海などからも産出される、エディアカラ紀の代表的なベンド生物のひとつ。同時にフィロズーン(Phyllozoon)の化石(写真なし)もありました。こちらもディッキンソニア(Dickinsonia tenuis)。南オーストラリア産。左右対称ではありません。この写真ではわかりませんが、中央の縦線から横方向に伸びる線が互い違いに出ています。また地衣類かもしれないという説もあるようです。(教養・文化シリーズ NHKスペシャル 生命大躍進より引用。)パルバンコリナ(Parvancorina minchami)。T字構造から左右対称かもしれないとされていますが、これはエディアカラ生物群としては珍しいです。節足動物の祖先と考えられたこともあります。次はカンブリア大爆発(5.4億年前~4.3億年前)。この時期、堅い外骨格を持つ生物が突如出現し、さらに主な動物の祖先がすべて出現。まさにエクスプロ―ジョンです!驚嘆すべきはエディアカラ生物群からわずか3000万年で眼や脳などを獲得したこと。特別展会場内にある、眼の獲得に関するロドプシン遺伝子の移動仮説の動画はとても興味深いものがあります。ぜひとも会場へ。展示においては、カナダのロッキー山脈のバージェス頁岩動物群、中国の雲南省のチェンジャン生物群の化石が盛りだくさんでした。アノマロカリス、オパビニア、ウィワクシア、ハルキゲニアなど、一見の価値ありです。個人的に注目した展示はピカイア(Pikaia gracilens)。カンブリア紀中期のバージェス頁岩動物群のひとつ。脊索を持っており(脊索動物)、脊椎動物の祖先にあたります。確かにナメクジウオのような外観です。メタスプリッギナ(Metaspriggina walcotti)。カンブリア紀中期のバージェス頁岩動物のひとつ。注目すべき点は眼、咽頭を持った脊椎動物である点。(頭部を含む破片とそのほかの体を含む破片に分かれています。)そして最古の脊椎動物と考えられているハイコウイクティス(Haikouichthys ercaicunensis)。カンブリア紀前期のチェンジャン生物群のひとつ。レプリカです。かなり小さく体長は2-3cmほど。眼がある軟骨性の頭骨を持ち、鰓らしい構造が確認できるとのことです。顎はありません。群れをつくっていた可能性が高いらしいです。ちなみに脊椎動物においては生命大躍進、ゲノムの4倍化が起こります。これにより多くの遺伝情報、新たな遺伝情報を伝えることが可能になりました。次はオルドビス紀(4億8500万年前~4億4300万年前)の展示。こちらはクセナサフス(Xenasaphus devexus)。オルドビス紀中期、ロシア産のアサフス科の三葉虫の化石です。カンブリア紀の三葉虫が平たいことに比べて、オルドビス紀の三葉虫は厚みがあり、盛り上がっています。説明には、まとまって産出することから群れて生活していたらしいと書かれてありました。ほかにもウミユリなどの化石の展示や大量絶滅についての展示がありました。シルル紀(4億4300万年前~4億1900万年前)の展示で印象的だったのは、ウミサソリ/アクチラムス(Acutiramus macrophthalmus)。シルル紀後期、アメリカのニューヨーク州産の化石です。写真は鋏角の部分。迫力ある体長2.2mの標本展示もあり、いっしょに写真を撮られている方がたくさんおられました。デボン紀(4億1900年前~3億5900年前)の展示で興味深かったのは三葉虫の化石。以下はそのごく一部です。パラホマロノータス(Parahomalonotus sp.)。デボン紀、モロッコ産の化石。ケイルスス科 属種不明(Cheiruridae gen. et sp. indet)。デボン紀、モロッコ産の化石。アカンソピゲ(Acanthopyge haueri)。デボン紀中期、モロッコ産の化石。この時期の三葉虫の化石を閲覧する上で、複眼(大きめのレンズを持つもの、小さめのレンズを持つもの)や突起・棘と言った構造に注目して見ていくとさらに面白いです。(ちなみに三葉虫のレンズはヒトデ類およびクモヒトデ類同様、方解石。)次は魚類の化石の展示です。魚類は大きく無顎類、板皮魚類、軟骨魚類、棘魚類、硬骨魚類に分けられ、硬骨魚類は条鰭類と肉鰭類に分けられます。シルル紀から白亜紀までの貴重な化石の展示に対して思わず感嘆・感激~。カンブリア大爆発のときにも思ったことではありますが、これだけの魚類の化石の展示だけでも特別展が開けそう~。それほどにおのおののセクションが充実しています。無顎類のロガネリア(Loganellia scotica)。シルル紀後期、イギリスのスコットランド産。顎がない魚です。体表には小さな鱗があり、胸鰭状の構造物があります。板皮類のコッコステウス(Coccosteus cuspidatus)。デボン紀、イギリスのスコットランド産。ダンクルオステウス同様、顎を持つ魚ですが、全長はダンクルオステウスと比べてかなり小さいです。軟骨魚類のクセナカントゥス(Xenachanthus sp.)。ペルム紀、ドイツ、メイゼンハイム産。淡水に生息していたらしいです。オルサカントゥス同様、頭部後方へ伸びる突起は特徴的です。棘魚類のアカントーデス(Acanthodes bronni)。ペルム紀前期、ドイツ、ラインラント=プファルツ州産。棘魚類のアカントーデス(Acanthodes bridgei)。石炭紀後期、アメリカ、カンザス州産。棘魚類の繁栄のピークはデボン紀。早くに顎を獲得した魚です。尾鰭を除く各鰭の前縁に棘を持っています。硬骨魚類の条鰭類、ケイロレピス(Cheirolepis canadensis)。デボン紀前期、カナダのケベック州産。同じく条鰭類のレピドーテス(Lepidotes minor)。ジュラ紀後期、イギリスのバーベック産。そして肉鰭類のユーステノプテロン(Eusthenopteron foordi)。デボン紀前期、カナダのケベック州産。両生類への進化において紹介されることが多い魚です。胸鰭には上腕骨、橈骨、尺骨があります。そしてデボン紀にはついに陸上への脊椎動物の進出が始まります。写真はアカントステガ(Acanthostega gunnari)の尾部。デンマークのグリーンランド産。レプリカです。F/F境界絶滅後のデボン紀後期に出現した初の両生類です。指は8本。橈骨が尺骨の2倍近くもあり、また関節が簡素とのこと。どうやら陸上での歩行には向いていない構造のようです。こちらはイクチオステガ(Ichthyostega sp.)の頭部。デボン紀後期、デンマークのグリーンランド産。レプリカ。アカントステガよりも大きな両生類。特徴的な太い、長い肋骨は内臓を保護し、陸上での体の支えになっていたのではないかと推察されています。そしていよいよ爬虫類の登場です。こちらは1対のメソサウルス(Mesosaurus tenuidens)の化石です。ペルム紀前期、ブラジル産。このメソサウルス(Mesosaurus tenuidens)の胎児がこちら。上:ペルム紀前期、ウルグアイ産の胎児化石のレプリカ。下:胎児の再現図。再現図と比べてみるとわかりやすいです。中央やや上の帯状のものが頭骨・下顎骨。またわかりにくいですが、中央やや左下の孤状のものが肋骨です。この化石には卵殻こそ確認できませんが、卵歯らしきものが見つかっており、卵殻があったと考えられています。また孵化直前の胚と考えられています。(ここで生命大躍進としての胚膜の誕生です。)石炭紀の後期に両生類から進化したグループとして単弓類がいます。単弓類とは眼窩後方の側頭窓という孔が片側に1つ、両側にあるグループです。哺乳類の祖先にあたり、ペルム紀にはこの単弓類が繁栄していました。展示されていた単弓類の一部がこちら。コティロリンクス(Cotylorhynchus romeri)。ペルム紀前期、アメリカ産。レプリカです。特徴は巨体と小さな頭であり、植物食だったと考えられています。そしてディメトロドン(Dimetrodon limbatus)。ペルム紀前期、アメリカのテキサス州産。レプリカ。個人的に好きなディメトロドンです。大きな頭骨と鋭い歯、見るからに肉食性ですね。背中にはエダフォサウルス同様、大きな帆のような構造があります。解剖学的に面白いのは、帆の骨が背骨の棘突起が伸びたものであること。帆には温度調節機能があったと推察されています。三畳紀前期の単弓類、獣弓目のリストロサウルス(Lystrosaurus georgi)。ロシア産。レプリカ。ペルム紀の大量絶滅を乗り越えて生き延びた4属のひとつ。(残りの3属は三畳紀の初めに見られたのみ。)化石は南極、アジア、ヨーロッパなどで見つかっており、超大陸パンゲアの存在、大陸移動説の根拠のひとつになっています。地上で恐竜が繁栄していたころ、哺乳類はひっそりと生き延びていました。ジュラマイア(Juramaia sinensis)です。ジュラ紀後期、中国の遼寧省産。レプリカ。胎盤を持つ最古の哺乳類とされています。(このときすでに胎盤形成にPou5f1およびCdx2遺伝子の関与あり?)ジュラマイア(Juramaia sinensis)の巣の模型。授乳して子育てをしている様子。ただし授乳様の行動はすでにキノドン類で行われていた可能性があります。こちらはアメリカ合衆国・コロラド州で見つかったK/Pg(白亜紀/古第三紀)境界層を含む岩石の断面です。断面下部の乳白色の帯状ラインが境界層です。白亜紀末の恐竜を含む大絶滅の原因として考えられている隕石衝突。地球上にはほとんど存在しないイリジウムですが、この境界地層からは高濃度のイリジウムが検出され、隕石衝突説を有力なものとしています。この大絶滅を生き延びた哺乳類が、恐竜が占めていたニッチを埋めるように放散しました。これら新世代の哺乳類の一部を紹介します。アイルラブス(Ailuravus macrurus)。始新世中期、ドイツのメッセル産。初期の齧歯類。パレオキロプテリクス(Palaeochiropteryxs tupaiodon)。始新世中期、ドイツのメッセル産。皮膜や体毛のあとまで残っている美しいコウモリの化石。ヒゴテリウム(Higotherium hypsodon)。始新世前期中期境界頃、熊本県宇土市産。裂歯類の右下顎骨の化石。歯冠の高い2本の臼歯が確認されます。メソヒップス(Mesohippus sp.)。始新世後期~漸新世前期、アメリカのサウスダコタ州産。初期のウマ類。前臼歯が臼歯化しはじめているものの、まだ低歯冠です。(進化するにつれて臼歯は高くなります。)このあと人類への道のセクションへ…。ヒトを含む霊長類の進化について興味深い展示がたくさんありました。こちらはプレシアダピス(Plesiadapis tricuspidens)の下顎骨の化石。暁新世後期、フランスのベル産。霊長類に似た大臼歯や小臼歯を持つとのことです。ただし特別展図録のXijun Niの記載によると、プレシアダピス類が霊長類の姉妹群であろうとする説にはいまだ議論が続いているとのことです。アルタニウス(Altanius orlovi)の上顎骨(上)および下顎骨(下)のレプリカ。始新世前期、モンゴル産。アルタイ山脈より見つかりました(名前の由来)。霊長類として最も古い化石のひとつ。アダピス類とオモミス類の共通の祖先に近い仲間のようです。そして今回注目のダーウィニウス・マシラエ(Darwinius masillae)。「イーダ」の愛称で知られています。このイーダの美しい化石がドイツのヘッセン州メッセルにて見つかりました。この化石のレプリカがこちら。始新世(4700万年前)。霊長類化石として完全な美しい標本です。当時のメッセルには火山があり、有毒ガスが立ち込め、水の中には酸素がほとんどなかったようです。そのため水に落ちた死骸の腐敗が遅延、すなわち分解を行う酸素を必要とする細菌が生存できなかったことがこの美しい奇跡の化石をつくったのだろうと考えられています。もちろんほかにも美しい化石がたくさん見つかっています。すでに新世代の哺乳類として掲載している、アイルラブスやパレオキロプテリクスがこれに当たります。パレオキロプテリクスといったコウモリ類、鳥類の化石はたくさん見つかっており、当時立ち込めていた有毒ガスを吸って絶命したと推察されています。ちなみにこのイーダがキツネザル類を含む原猿類なのか、真猿類なのかについてしばらくの間議論されていましたが、現在はほぼ前者であると考えられています。人類進化の道すじのセクション。人類の進化についての説明、充実した化石・レプリカの展示です。その一部を紹介。猿人、サヘラントロプス・チャデンシス(Sahelanthropus tchadensis)。中新世(700万年前)、チャドのトロスメナラ産。レプリカ。人類化石のなかで最も古いとされています。「サヘラン」とはサハラ砂漠の南と言う意味らしいです。今回いろいろな角度から観察させていただきました。頭蓋を見ると脳は小さそうですが、大きな犬歯を確認することはできず、大後頭孔の位置は斜め下方から確認すると中央付近でした。人類(猿人)としての判断はこのあたりがポイントのようです。原人、ホモ・ハビリス(Homo habilis)。更新世(190万年前)、ケニアの東トゥルカナ産。レプリカ。アウストラロピテクス属から進化した2系統のうちのひとつ、ホモ属の初期段階です。歯が小さくなっています。ホモ属では脳の大型化が始まります。ちなみに更新世(170万年前)のホモ・エレクトス(Homo erectus)では脳容量がかなり増大しました。ホモ・エレクトスよりさらに進化した旧人、ホモ・ハイデルベルゲンシス(Homo heidelbergensis)。更新世(43万年前)、スペインのアタプエルカ産。レプリカ。もとはドイツのハイデルベルグ付近で見つかった下顎骨の化石人類につけられた名前です。脳容量はさらに大きくなり、1100~1400ccほど。火を使うようになりました。こちらは旧人、ホモ・ネアンデルターレンシス(Homo neanderthalensis)。更新世(6万年前)、イスラエルのアムッド産。レプリカ。いわゆるネアンデルタール人です。脳容量は1500~1600ccと増大。石器などで狩猟を行い、死者の埋葬を行っていたようです。ただし石器はさらに以前の260万年前から使用されており、アフリカのオルドヴァイ渓谷などで見つかっています。そして私たち新人、ホモ・サピエンス(Homo sapiens)です。完新世(5000年前)、日本の宮野貝塚産の縄文時代人の実物化石。ホモ・サピエンスは20万年前にアフリカで生まれたとされています。1つ前のネアンデルタール人の頭蓋骨と比較すると、眼窩前上縁の膨隆が目立ちません。頬骨の突出も目立たないような?(気のせい?)とここで終わりではなく、最後には「受け継がれたDNA ~ヒトゲノムに刻まれた進化の痕跡~」のパネル展示とビデオ上映がありました。学生のころから勉強せざるを得なかった遺伝子。メンデルから始まり、ワトソンとクリックによるDNAの解明、そしてその後もいろいろと…。いまだ仕事上、何らかの形で関わりのある遺伝子です。パネルでは現在解明されている遺伝子情報についての掲示があり、既知の知見以外の新たな知見から多くのことを学ばせていただきました。そして気の遠くなるような歳月ををかけて受け継がれ進化した遺伝子情報には、驚きと不思議を隠せませんでした。今回、生命の誕生からヒト(ホモ・サピエンス)までの進化をテーマに、DNAという角度から経時的、そして段階的に化石を主体に展示および解説がなされていました。一般的にDNAというとなんだかとっつき難い印象があるものの、解説は親切丁寧でわかりやすく、おのおののDNAの進化に関する化石の展示が充実しており、心に残る、非常に充実した特別展だと感じました。長々とブログに掲載いたしましたが、それでも展示のごく一部にしかすぎず、ほかにも多くの見どころがあることには違いありません。また会場で売られている特別展の図録の美しい写真と解説を読むことで、個人的にもう一度行ってみたい気になりました。今後、これらを含む遺伝子情報の解明が、遺伝子治療などの多くの分野で役立つことを期待しながら、今回の特別展の備忘録を終わりにしたいと思います。
2016年04月29日
-

ミカドウニ、少しだけ移動した
ミカドウニ(Goniocidaris mikado)です。横向きになってようやく観察することができた管足~♪本当に生きているのかとさえ思っていただけに管足を確認できたことはうれしいことです。しかもほんの少しだけ移動しました。かなりゆっくりでしたけれど…。~~~~~また少し寒くなりました。困ります。やむを得ず、室温でトリートメントしていた個体に対して、サーモスタット、ヒーターセッティングを行いました。
2016年04月28日
-
つぶやき:最近のこと
1.少し前に来た「海洋と生物」をちらほらと斜め読み…。 のつもりでしたが、一部熟読。 興味ある深海関連のことが特集だったから仕方ないですね(笑)。2.以前よりジェネリック薬品への変更が進行中。 変更を前もってアナウンスしてくれるのはいいのですが、 それでも何の後発品かわからないものが徐々に増加…(汗)。 調べなくては。(前にも書いたような?)3.神経伝導路の二次変性について、 過去論文のまとまった知見が得られ、一部の疑問が解決しました。
2016年04月27日
-

深海よりウミエラの仲間
深海生体の一時的なキープ槽より取り出したウミエラの仲間。手袋をしているのはウミエラに対してではなく、いろいろいるのでそっちのほうを警戒。例えばウミケムシ類とかね。処分してからは見ないけれど、念のため。それに手袋で間接的に生体を触ることで、皮膚温との温度差を緩和。手袋内にも海水が入っているのでかなり間接的だと思います。ちなみに水温は13℃ほど。このウミエラは今後別の場所へ移動予定です。今回はここにいるほかの生物を調べることが目的でした。いろいろいるのですが、実は把握できていません…(汗)。怠慢な自分にちょっと反省です。
2016年04月26日
-
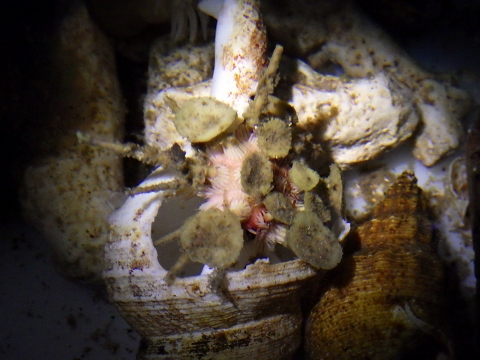
ミカドウニ、その後
ミカドウニ(Goniocidaris mikado)です。水槽へ入れてからしばらく経ちました。当初、棘関節部分で浮いていた2本の棘は外れてしまいましたが、それ以外の棘は問題ないようです。それよりも本当に生きているのかどうか、不安になるほどに動かないです。それでときどき取り出して、腐敗臭がしないかどうか確認したりしましたけれど問題なし。翌日には居る場所が変わっていることがあり、移動はしているようですが、未だ移動している瞬間を確認できておりません。かなり静的な生き物。興味は尽きませんが、状態を把握しにくいのが難点(汗)。(棘がぼろぼろと取れはじめたらダメなんでしょうが…。)
2016年04月25日
-

再生した第2触角
オオコシオリエビ(Cervimunida princeps)です。無事脱皮を終えました。当初より欠損していた両側の第2触角は再生されました。めでたし、めでたし~♪ところで脱皮殻を食べるのだろうか?硬そうだけれど…。しばらく放置して様子を見ようと思います。
2016年04月24日
-

小競り合い?
ツノガイに入った深海性ヤドカリ。ピンボケすみません。左下のヤドカリが右上のヤドカリの歩脚をつかんで離さないようです。ときどき見られる小競り合い?でもこれが原因で歩脚がなくなってしまったことは今のところないです。
2016年04月23日
-

数cmにおよぶ脱腸、その後の腸管壊死から回復した個体
以前より飼育している沖縄採集の深海魚バラハナダイ。何匹かはすでに掲載していますが、今回の1匹は初登場です。深海魚専門のUさんといっしょに採集したものの、鰾破裂、数cmにおよぶ脱腸状態であった個体。鰾破裂は何とかなることが多いですが、後者は非常に厄介です。脱出腸管の環納はまず不可能で壊死に陥ることはほぼ必須。壊死には虚血・感染・損傷が影響しているのかもしれませんが、今のところどうしようもない状態には違いありません。壊死腸管が脱落するまで生存したとしても、創部の易感染・易出血状態や脱落後の治癒過程における癒着性の腸閉塞など、危惧する点は多いと思っています。そういった点において今回生存しえたこの1匹は貴重であり、治療なされたUさんには誠に感謝いたしております。現在摂餌・排泄はともに良好。尾鰭下葉はやや小さいですが、Uさんいわく、他のバラハナダイに攻撃されたとのことでした。5か月ほど経過しましたが、今のところこれ以上の再生はないようです。仕方ないですが、これ以外には特に問題がないので安心しています。
2016年04月22日
-
つぶやき:最近のちょっとしたこと
1.某薬剤師さんから電話あり。どうやら引っ越したらしい。 家が近くなったので、今度行ってみるか。2.学会にて製薬会社、医療機器メーカーの展示ブースへ行ってみた。 ここにいると言っていた某MRがいない。残念。3.水槽クーラーのひとつの調子が良くない。 温度センサーがダメになっている可能性あり。 とりあえず予備クーラーと交換。 機器トラブルによる生体への影響を考えるとゾッとする。
2016年04月21日
-

右胸鰭のcystic lesion
カリブ海の深場からやってきたディープウォータースコーピオンフィッシュ(商品名)。以前より飼育を続けており、掲載はかなり久々。前回の掲載は8か月以上も前でした。その間ネタがなかったからねぇ…(汗)。ご覧のようにこのリフジウム水槽には活性炭の入った袋を底面にそのまま放置しています。なんともずさん…(汗)。さて今回この魚の右胸鰭にちょっとした異常が見つかったので掲載。見つかったのはオタマジャクシのような形をしたcystic lesion。隣接する軟条の腫大や損傷はなく、鰭膜の病変なのかもしれませんが、本当のところはよくわからないです。病変と周囲組織との境界は明瞭で、内部に明らかなsolid partはないように見えます。個人的にはbenign lesionの可能性が高いと思うので、このまま経過観察しようと思っています。でももし増大傾向があれば、そのときは何からの処置を行うべきでしょうね。
2016年04月20日
-

イソギンチャクに群がるヨロイウミグモ、体液を吸うらしい
ヨロイウミグモ(Pycnogonum tenue)です。ホムラspのところへやってきてしばらく経ちました。実は以前より飼育したいと考えていたヨロイウミグモ。ついに念願がかないました。イソギンチャクの体液を吸うらしいので、いっしょに深海性のイソギンチャク類を入れています。イソギンチャクは開いていませんね…。群がりすぎかな(苦笑)。
2016年04月19日
-

またもや見つかる変なやつ
深海の沈木にいつも見られるサイフォン。石灰質の棲管に入っているようです。フナクイムシ類なのかな。こちらもいつも見られる沈木の穴の中のサイフォン。キクイガイ類だと思うのですが。いずれにせよ、沈木を割って確かめたい衝動にかられます(笑)。餌のにおいにつられて出てくる環形動物類。苦手な人は多いと思います。そして今回新たに見つかった変なやつ。これも環形動物なのかな?いろいろ見つかって楽しいものです~♪~~~~~学会で恩師の講演を聞いたり、興味ある発表を聞いたり…。勉強になりますねぇ~。学会の会場には専門書がたくさん売られていたので、数冊購入。持ち帰るには重たかったので郵送していただきました。早速、仕事で活用しています。
2016年04月18日
-

関東へ
少し前に新幹線に乗って関東方面へ出かけました。新幹線の車窓からの眺め。このあたりはまだ元気なんですが…。ここまで来ると睡魔に勝てずに寝ていることが多いです。でも今回は起きていました。私用のために丸1日がつぶれてしまいました。仕様のない私用(笑)。~~~~~新たなリフジウム水槽をセッティング。同時にトリートメントを終えた魚を入れて現在遮光中。遮光しないとなかなか落ち着かないんだよねぇ、この魚は…。
2016年04月17日
-

いるときでよかった~
リュウグウハゼ(Pterogobius zacalles)です。少し前にとび出しました。蓋をしているのですが、簡易にのせていただけで隙間があった可能性あり。でもいるときでよかった~。~~~~~ナメダンゴ(メス)が残念なことにお亡くなりになりました。飼育期間はおよそ3年1か月でした。
2016年04月16日
-

まめな給餌が続くなり
深海性ホヤに付着する瓦礫の中に隠れていた樹手目のナマコ類。触手で水中の微小なエサを摂取する、いわゆる濾過摂食を行うナマコです。まめな給餌が続きます。そしてある程度の水流が必要。これらを怠ると触手を伸ばさなくなりますが、水流だけでも伸ばしていることは多いです。ちょっと厄介、でも楽しかったりして(笑)。
2016年04月15日
-

パープルリーフロブスター:脱皮後の奇異な状況
今まで何度か脱皮を行ったパープルリーフロブスター(Enoplometopus debelius)。今回の脱皮後の状況は少し奇異でした。過去数回の脱皮後の状況と異なる点は、1.脱皮殻をほとんど食べなかった。2.よく食べていたオキアミをほとんど食べなくなった。3.塩ビ管からほとんど出なくなった。(写真は状態観察のため塩ビ管から出した際に撮影したもの。)こんなところ。いずれにせよ、脱皮後数日経っても積極的な探餌行動・摂餌行動は見られず、与えたオキアミをほんの少し摂餌するのみで、大半を残していました。そこで気になったことと言えば、1.脱皮のマイナーな失敗。2.感染症の罹患。3.何らかのストレス状態。4.嗜好の変化など。見た目脱皮を成功させていたのですが、わからない部分で失敗があったのではないかと危惧しました。感染症などの病気ということも。飼育環境に対するストレス発生ということも。でも今までと同じ飼育環境だし…。4の嗜好の変化。まず最初にこれに対して餌をいろいろ与えてみて、確認してみることにしました。キビナゴの身、キビナゴの臓器類、オキアミ、乾燥クリル、練り餌、粒餌、フレークフードなど。この中で比較的よく食べたのがキビナゴの臓器類でした。次にミネラル+ビタミンの添加を行った練り餌も食べることを確認しました。嗜好の変化、何らかの栄養素の要求状態なのかなぁ?とりあえずの現況はこのような感じ。これから続けて試行錯誤していこうと思います。
2016年04月14日
-

カセミミズのその後
低温水槽で飼育中のカセミミズ(Epimenia verrucosa)です。小さなトサカのある水槽に一時的に入れたものの、その後いろいろあって現在の低温水槽で観察しています。ここには多数のクラゲのポリプが付着しています。小さなエフィラらしきものも探すと簡単に見つかります。そういった点で個人的にとても面白い水槽~♪さて肝心のカセミミズですが、貝殻に巻きついたり、壁面に付いていたり…。じっとはしているものの、ときどき場所の変化があります。(食べないと思うけれど)クラゲのポリプを食べたら面白いなぁ~と思ったりもしましたが、食べているところは確認できていません。体色の変化は気になるところ。斑点は黄色じゃなくなってしまいました。あと途中から同居しているジンゴロウヤドカリはカセミミズに対して全く興味がないようです。襲うようなことがあれば、即刻退去だったんですけれどねぇ~。以上、最近のカセミミズでした。
2016年04月13日
-

排泄物の分解が遅いなり~
Antennarius biocellatusです。比重1.008での飼育を行っている汽水性のカエルアンコウの仲間です。現在に至るまで見た目の異常はなく、摂餌・排泄ともに良好。ここまではいいのですが、ちょっとだけ悩み事が…。それは排泄物の分解が非常に遅いこと。濾過槽を確認すると明らかです。視認する上で水槽内に微生物はほとんどいないからなぁ~。たまに線虫らしきものがいるくらい。その上バクテリアの添加を行っていないことが一因。まぁこのあたりが原因なんでしょうね。ただバクテリアに関しては少々慎重です。汽水を好む遅発育菌のMycobacterium marinumが厄介…。この点においてバクテリアよりも殺菌を優先させてしまいがち…。結局、排泄物の分解が遅くなるのは当然の結果なのかな…(汗)。
2016年04月12日
-

ピンクの眼をしたヨコエビの脱皮
以前に掲載した深海性のホヤ(水深300m)に付着していたヨコエビの仲間のことです。その後ホヤといっしょに観察を継続。特にこれと言ったエサを与えるでもなく、海水に溶かした粉末状のエサ+αをホヤに給餌する際にいっしょに食べてくれていたらいいなぁ~と考えていました。さてそのヨコエビの仲間ですが、本日、脱皮していました。5mmほどの小さなヨコエビの仲間。いろいろ付着したホヤの表面につかまったままの状態で動かず…。ピンク色の眼がとても目立ちます。隣には脱皮殻がありました。こういった小さな生き物の頑張りには何らかの感動を覚えます。これからも元気でいてくれよ~。
2016年04月11日
-

新たな深海性イソギンチャクを発見
深海生体と深海の沈木のみを入れている水槽の底面に、いつの間にか小さなイソギンチャクが付着していました~♪このことに気付いたのは数日前のこと。そのときに取り出してケースへ移し、飼育を試みています。写真左のイソギンチャクの口盤の直径はだいたい5mmほど。写真右のイソギンチャクはさらに小さく、半分ほどしかありません。(あまり大きくならない種なのかなぁ?)深海性のイソギンチャクとして興味深いものがあるので、このまましばらくの間、飼育したいと思います。
2016年04月10日
-

素晴らしき出会いに感激
ミカドウニ(Goniocidaris mikado)です。熊野灘で採集された深海性のウニ。特徴的な皿のような棘、一度見ると忘れられないインパクトがあります。一部の棘が棘関節部分で浮いているように見え、深海採集のダメージがあると考えられるものの、それ以外には特に目立ったところはなく、比較的良い状態です。もちろん今回が初飼育。これからが楽しみです。ちなみにミカドウニを含むキダリス類は三畳紀からほとんど変化をしていない生きている化石みたいなウニです。殻板やランタンの構造などには特徴があり、個人的には非常に興味深いものがあります。
2016年04月09日
-

今回はちょっと長め~
ハシキンメ(Gephyroberyx japonicus)のこと。1匹のハシキンメが摂餌しなくなってからおよそ2か月が経過。少し心配でしたが、最近になり再び食べ始めました。時々食べなくなることはありますが、今回はちょっと長めでしたね。その間、エサに対する興味は落ちていないようでした。エサに対して近寄ってはいたのですが、それ止まり…。最後の食いつきがなかなか見られませんでした。現在は食べているので安心していますが、いまだに長期にわたり食べないときとの違いがわかりません…(汗)。
2016年04月08日
-

ヤドカリ、貝を食す
写真は深海生体とともにやってきたヤドカリ。以前に掲載し、それ以後1回脱皮を行っています。まだ名前を調べていませんが、深海性ヤドカリとして気にはなるところ。写真右の深海性のウニはPrionechinus forbesianus。もう何度か掲載しています。沈木生体として沈木とともに飼育中です。ほかに沈木生体としては貝類が多く、それらも同時に飼育しているのですが、その一部の貝をヤドカリが食べてしまいました。右が飼育中の貝で、左が食べられた後。ヤドカリにとっては格好のエサだったんですね…。現在(最初の)ヤドカリの移動先を考慮中です。
2016年04月07日
-

ツノガイに入った深海性ヤドカリの飼育経過
ツノガイに入った深海性ヤドカリ。この日はオキアミ小片を与えました。摂餌しています。結構いろいろなエサを与えていますが、今のところよく食べています。今回いつまで飼育できるかわかりませんが、前回飼育時と比べて比較的順調な印象があります。前回飼育時と異なる点と言えば…。1.水温が1℃ほど低い。2.最初から底砂をひいている。3.水流がほとんどない。こんなところ。(2は当然すべきことだったんですけれどね。)しばらくこの状態で飼育を続けてみようと思います。
2016年04月06日
-

指を数か所刺されました
ライブロックをデッドロックにしてから使用することが多くなり、水槽ではほとんど見ることがなくなったウミケムシ類。最近では沈木の中にいたウミケムシ類らしきものを見たぐらいでそれ以前はほとんど見かけず、そして刺されることはなくなっていました。でも沈木にそれらしきものがいたんですね。排除したあと、すっかり忘れていました。沈木には剛毛が付着し、残存している可能性が十分あるわけで…。結果…。昨日、不注意にも沈木を素手で触っていて、指を数か所刺されました。写真は指に刺さっている剛毛(?)らしきものの。かなり拡大していますが、実際は光を当ててじっくり観察しないとわかりにくいです。チクチクと何とも言えない痛みがあります。ピンセットでつまむと簡単に折れるので、折れないように爪でそっとつまんで引き抜きました。一部はセロハンテープで横から寝かすようにして貼り付け、除去しました。処置後、局所の腫脹・発赤・疼痛はなかったので、薬剤塗布を行わず、放置しています。もちろん全身症状もなし。(あればcase reportとして興味深い、いやそんなことを言ってられんかも(苦笑)。)写真の剛毛(?)らしきものはウミケムシ類のものなのかどうかわかりませんが、以後注意しないといけませんね。
2016年04月05日
-

Deep water Xylophagaidae:深海の沈木から
熊野灘の水深350mの沈木の中から見つかったキクイガイ(二枚貝)の仲間です。見たのは今回が初めて。このように沈木外に出ていることはなく、普段は沈木の中に穿孔した状態で生息しています。今回、写真左側部分のsiphonだけがかろうじて沈木の小さな穴の中に見えていました。それで慎重に沈木を割って取り出しました。せっかくだから撮影。背側から。前方から。そして側面。殻長は11mmほど。siphonは10mm近く伸びていることがあります。沈木を観察すると同様のsiphonらしきものが穴の中にあるような?ほかにも生息しているのかもしませんね。また、キクイガイの仲間だけでなく、フナクイムシの仲間らしき石灰質の棲管とsiphonもあります。まだまだ楽しめそうな沈木観察です。今後にも期待~。
2016年04月04日
-

落ち込んだ彼女を救え
ユビナガワモンヤドカリ(Ciliopagurus krempfi)です。彼女は大変な状況なんです。そう、彼女なんですね。昨年脱皮した殻の生殖孔の位置からメスであることがわかっています。話を戻して…。飼育生体の増加などにより移動させられたユビナガワモンヤドカリ。深海の沈木生体がいる場所に一時的に移動させたものの、仕切り板と沈木の間に落ち込んでしまい、出られない状況になっていました。宿貝の突起が引っ掛かって、上にも下にも動けない状況。ピンセットで取り出しましたが、引っ掛かりがなかなか取れず、少々苦労しました。別の場所への移動を考慮すべきかもしれませんね。
2016年04月03日
-

たまには水槽掃除を…
ヒメセミエビ(Chelarctus cultrifer)の飼育開始を初めて1年1ヶ月が経過しました。その間脱皮は1回。これ以外に目立ったイベントはなく、今回久々の登場です。4月2日はこのヒメセミエビの飼育水槽を掃除し、水換え、そしてレイアウトの変更を行いました。写真はその数時間後の給餌のときに撮影したもの。エサにつられて出てきました。かわいいものです~♪ほかにも掃除を行いたい水槽があるのですが、ややさぼり気味…。あと病気発生リスクのことを考えると魚水槽はやりづらいものがあります。
2016年04月02日
-

深海魚ヒメの仲間の飼育
沖縄の深海220mから採集されたヒメの仲間です。(個人的にはハタタテヒメの雌、でもヒメの可能性も…。)飼育開始から半年が過ぎ、もう数日で7か月になります。以前に飼育していたハタタテヒメの雄の飼育期間を超えることができました。前回のハタタテヒメの飼育経験を活かして臨んだ今回の飼育。今のところ摂餌・排泄となかなかいい感じかなぁ~と思っています。しかしながらまったく問題点がないわけではありません。以前にも記載したことがありますが、腹鰭鰭条先端の底面に接する部分が白っぽくなり、棍棒状に太くなっています。右腹鰭のほうがやや目立ちます。アクリル水槽底面との接触による影響かも…。イリドウイルス感染の関与かも…。いろいろ考えています。ちなみに混泳しているバラハナダイとの相性に関してはまったく問題ありません。ピンボケすみません。いつものことです。
2016年04月01日
全30件 (30件中 1-30件目)
1
-
-

- (o>ロ<)o多頭飼いのダックス大好…
- さとにわ耕園へお散歩に行ってきまし…
- (2026-05-20 13:54:55)
-
-
-

- ポメラニアンとの毎日
- 💝【メルカリ】宛名シール 差出人シ…
- (2024-07-04 08:35:07)
-




